中央研究院 生物化學研究所
中央研究院 生物化學研究所
中央研究院 生物化學研究所
中央研究院 生物化學研究所
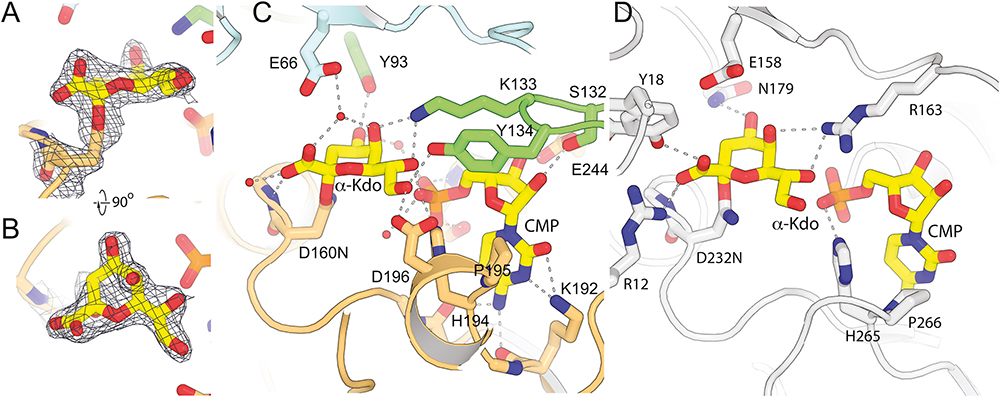
KpsC is a dual-module glycosyltransferase (GT) essential for "group 2" capsular polysaccharide biosynthesis in Escherichia coli and other Gram-negative pathogens. Capsules are vital virulence determinants in high-profile pathogens, making KpsC a viable target for intervention with small-molecule therapeutic inhibitors. Inhibitor development can be facilitated by understanding the mechanism of the target enzyme. Two separate GT modules in KpsC transfer 3-deoxy-β-d-manno-oct-2-ulosonic acid (β-Kdo) from cytidine-5'-monophospho-β-Kdo donor, to a glycolipid acceptor. The N-terminal and C-terminal modules add alternating Kdo residues with β-(2→4) and β-(2→7) linkages, respectively, generating a conserved oligosaccharide core that is further glycosylated to produce diverse capsule structures. KpsC is a retaining GT, which retains the donor anomeric carbon stereochemistry. Retaining GTs typically use an SNi (substitution nucleophilic internal return) mechanism, but recent studies with WbbB, a retaining β-Kdo GT distantly related to KpsC, strongly suggest that this enzyme uses an alternative double-displacement mechanism. Based on the formation of covalent adducts with Kdo, identified here by mass spectrometry and X-ray crystallography, we determined that catalytically important active site residues are conserved in WbbB and KpsC, suggesting a shared double-displacement mechanism. Additional crystal structures and biochemical experiments revealed the acceptor binding mode of the β-(2→4)-Kdo transferase module and demonstrated that acceptor recognition (and therefore linkage specificity) is conferred solely by the N-terminal α/β domain of each GT module. Finally, an Alphafold model provided insight into organization of the modules and a C-terminal membrane anchoring region. Altogether, we identified key structural and mechanistic elements providing a foundation for targeting KpsC.